In our last issue, we explored the physiology of digestion in the small intestine and started our discussion of nutrient absorption. In this issue, we conclude that discussion. Effectively, this is the heart and soul of our entire series on the digestive tract. Ultimately, everything that happens in the digestive tract is designed to get nutrients into the bloodstream. The final step in the process, absorption, is in many ways the most fascinating part of the discussion. Stomach acid unwinding proteins and pepsin breaking them down — that’s simple stuff. How the body actually recognizes amino acids and peptides and then transports them across the wall of the small intestine, that’s remarkably complex and fascinating…and important to understand in terms of optimizing your nutritional uptake and, ultimately, your health.
Note: this is a fairly technical discussion. However, my goal is to make sure you understand enough of it so that:
- You are never overwhelmed by the technical for very long.
- You walk away with an overall understanding of how nutrients are absorbed in the small intestine.
Absorption
As we discussed in our newsletter on the anatomy of the small intestine, virtually all nutrients, including all amino acids and sugars, enter the body by crossing the enterocytes (the absorptive cells found in the small intestine) that make up the epithelium layer that covers each and every villi (the hair-like extensions that project from the wall of the small intestine). There are two routes by which molecules make their way from the small intestine into the bloodstream:
- The transcellular route — across the plasma membrane of the enterocytes.
- The paracellular route — across tight junctions between the enterocytes.
For the most part, the tight junctions of the paracellular route are impermeable to large organic molecules such as dietary amino acids and glucose. Those types of molecules are transported exclusively by the transcellular route. Transcellular absorption of nutrients can take place by active transport or by diffusion. Active transport involves the expenditure of body energy, whereas diffusion occurs simply through random molecular movement and, therefore, without the use of body energy. Water for example, is transported through the intestinal mucosa by diffusion (isosmotic absorption); on the other hand, the absorption of amino acids and sugars involves active transport. This is one of the main reasons that eating a large meal can put you to sleep. You literally exhaust your body digesting and absorbing nutrients — until down the road, those same nutrients ultimately make their way into your body’s individual cells, thus, once again energizing you. Depending on the food you eat, you gain on the exchange — deriving more energy as the cells absorb the nutrients than was lost in digesting those nutrients and getting them into the bloodstream.
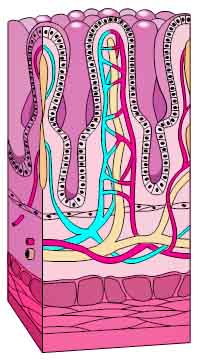 In any case, after passing through the epithelium into the villi, most of these molecules then cross over into the capillary network found inside each villus, thus making their way into the bloodstream. Fats, as we discussed when exploring the anatomy of the small intestine, behave differently. Instead of diffusing into the capillaries, they make their way into the lacteals, the lymphatic vessels present in each villus. From there, they drain from the intestine and rapidly flow through the lymphatic system and ultimately into the bloodstream by way of the thoracic duct.
In any case, after passing through the epithelium into the villi, most of these molecules then cross over into the capillary network found inside each villus, thus making their way into the bloodstream. Fats, as we discussed when exploring the anatomy of the small intestine, behave differently. Instead of diffusing into the capillaries, they make their way into the lacteals, the lymphatic vessels present in each villus. From there, they drain from the intestine and rapidly flow through the lymphatic system and ultimately into the bloodstream by way of the thoracic duct.
The process of crossing the epithelium into the villus, however, is not simple. In fact, the process varies for each nutrient. Or to put it another way, the epithelial tissue covering the villi is not uniform throughout the small intestine — or for that matter, from top to bottom in a single villus. Individual epithelial cells vary in both their makeup and functionality. In fact, each villus has a multitude of different receptor sites, specific for each nutrient. Each type of protein fragment and each type of carbohydrate fraction has its own particular receptor site it uses for absorption. In addition, as mentioned earlier, some nutrients diffuse through the spaces between the epithelial cells (the paracellular route) — spaces that vary throughout the intestinal tract, which has a significant impact on permeability. This becomes particularly important when we talk about the absorption of supplemental proteolytic enzymes (which are protein molecules). Unlike food proteins, proteolytic enzymes can actually use the larger spaces between cells to transport themselves out of the small intestine.
The bottom line is that as chyme (the mixture of broken down food and digestive juices) travels through the small intestine, it is exposed to a wide variety of absorption sites, each with very different characteristics. These absorption/receptor sites differ in the number and type of transporter molecules found in the plasma membranes of each individual cell. And once again, keep in mind, each individual villus is comprised of multiple enterocytes…each with a multitude of receptor sites. In other words, there are a vast number of receptor sites in the small intestine.
The chemistry of absorption
The key to the absorption of most nutrients in the small intestine is the electrochemical pump powered by electrolytes (primarily sodium) which works across the epithelial cell boundary of the villi. In fact, this is not unique to cells in the small intestine. Every single cell in the body is required to maintain a low concentration of sodium inside the cell (with a correspondingly high concentration of sodium outside the cell), which is required for the movement of nutrients into the cell and waste out of the cell. Correspondingly, potassium levels tend to be high inside cells and low in the areas just outside them. In addition, the sodium pump requires the presence of a large number of Na+/K+ ATPases (ATP enzymes) to regulate and power the reaction. This means that the cells of the body require the expenditure of energy (in the form of ATP) to power the sodium pump. The purpose of the sodium pump is to pull nutrients into the cell as sodium flows in and move waste out of the cell as potassium moves out. With that said, it’s now time to bite the bullet and get specific as to how nutrients move in and out of cells.
Every cell in the small intestine has three types of gateways that combine to move nutrients in and waste out.
- The actual sodium pump that is used to move sodium into the cell and potassium out of it. It carries three sodium ions into the cell and two potassium ions out with each action of the pump. (Don’t panic; we’ll explain this in more detail in a moment.)
- The leak channels for both potassium and sodium. If sodium continually moved into cells and potassium out, then in short order the cell would become electrically unbalanced. To help maintain the electrical balance of the cell, there are sodium and potassium ion “leak” channels in the membrane of each cell. These channels are normally closed, but even when closed, they “leak”, allowing accumulated sodium ions to leak back out of the cell and excess potassium ions to leak back in, as needed down their respective concentration gradients. In other words, the leak channels work in conjunction with the sodium pump, and are used to maintain the electrical differential that drives the pump. This is known as the cell membrane potential.
- The receptor sites make use of this electrical potential to carry nutrients (specific to each receptor site) into each cell. Let me repeat that one more time: each receptor site is specific to a particular nutrient. One receptor site transports glucose. Another site transports a specific type of amino acid. And so on. (A little later, we will discuss exactly how this works.)
To summarize, there are three types of gateways. The first two gateways are specific to the sodium pump and are used to maximize the potential of the cell to absorb nutrients. The third gateway is specific to pulling nutrients into the cell. Here’s a clean explanation of how it works.
By the way, there are approximately 150,000 sodium pumps per small intestinal enterocyte (cell). Each single cell is thus able to transport about 4.5 billion sodium ions into each cell per minute — along with accompanying nutrients. So with that in mind, let’s explore these specialized means of absorption in some detail:
Carbohydrates
Most dietary carbohydrates (even most simple sugars such as sucrose and lactose) cannot be absorbed in the intestinal tract. The monosaccharides (glucose and galactose), on the other hand, are actively transported with sodium. Monosaccharides, however, are only rarely found in normal diets. Rather, as described in Part 1 of our discussion of the Physiology of the Small Intestine, they are derived by enzymatic digestion of more complex carbohydrates in the small intestine. In summary, glucose and galactose are taken into receptor sites found on the villi by co-transport with sodium using the same transporter.
Now, for the briefest of moments, let’s get technical. (Hang in there; it’s actually understandable.)
The specific transporter molecule that carries glucose and galactose into the absorbing cell on the intestinal wall is SGLUT-1, also known as the sodium-dependent hexose transporter. This molecule will only transport the combination of a glucose and sodium ion into the cell together; it will not transport either molecule alone.
It works as follows:
 The transporter molecule is initially oriented facing into the small intestine. At this point, it can only bind sodium – not glucose.
The transporter molecule is initially oriented facing into the small intestine. At this point, it can only bind sodium – not glucose.- The act of binding sodium inside the transporter molecule triggers the opening of the glucose-binding pocket.
- This causes glucose found in the small intestine to also bind inside the transporter cell. The binding of the glucose molecule triggers the transporter molecule to reorient so that the pockets holding sodium and glucose are moved so that they face inside the cell.
- The sodium now moves off into the cell’s cytoplasm, which triggers the glucose to also unbind and move off into the cytoplasm.
- The emptying of the transporter molecule triggers it to reorient back to its original, outward-facing position. And the cycle starts again.
- The transport of galactose works in exactly the same way.
Once inside the enterocyte, glucose, galactose and fructose are transported out of the cell through another hexose transporter called GLUT-2 and on into capillaries that are found within each villus.
As we’ve already discussed, this is called active transport because it requires the use of ATP and requires the expenditure of some energy both for pulling the sugar molecules into the enterocyte, and then on out of the cell into the bloodstream. However, some time later, after using the sugars to power the body’s cells, the end result is a net gain of energy.
Fructose, of course, is the other simple sugar readily absorbed in the small intestine. The transport of fructose, though, involves an entirely different process. It is absorbed through something called facilitated diffusion (facilitated by Glut5) and requires no added energy (ATP) to cross into the bloodstream. The ability of fructose to be absorbed so easily into the system is indicative of its high reactivity in the body — and therefore also indicative of some of the problems it can present when consumed in a “pure” form such as high fructose corn syrup. When bound with fruit fiber, it behaves differently. It breaks down more slowly and is absorbed more slowly — thus presenting fewer problems.
As we mentioned earlier, the receptor sites for sugars are specific for sugars. This allows for an interesting option. Certain forms of fiber (which are also carbohydrates) can actually fill these receptor sites making them unavailable for use by the sugars for about an hour. Now, although these fibers can fill the sites, they are not transported into the cell. Instead, they occupy the site for up to an hour (again making those sites unavailable to any sugars for that period of time) until they are eventually rejected by the gateway and move out of the receptor site, then on down the digestive tract and out through the large bowel. Why is this important? Because the use of a sugar metabolic enhancement formula based on these fibers can modulate sugar uptake — slowing down and evening out the absorption of sugar — thus helping to avoid insulin spikes. The health benefits can be profound.
Proteins
After digestion, the proteins consumed in our food have been broken down into single amino acids, dipeptides, and tripeptides. These protein “pieces” are actively transported across the duodenum and jejunum. In fact, the mechanism by which amino acids are absorbed is virtually identical to that of monosaccharides, but takes place in different receptor sites. Amino acids are transported by sodium through nutrient gateways built into the cell walls of enterocytes. Dipeptides and tripeptides, on the other hand, are transported in a similar manner, but with hydrogen, not sodium, as the transporter. Again, since we’re talking about active transport involving the use of ATP, varying amounts of energy are required in the absorption of proteins.
It should be noted that as with carbohydrates, the transporter receptor sites are specific to amino acids and specific to different types of amino acids. In fact, there are several sodium-dependent amino acid transporters — including one each for acidic, basic, and neutral amino acids. Once again, these transporters bind their specific amino acids only after binding sodium. The fully loaded transporter then dumps sodium and the amino acid into the cell’s cytoplasm, followed by its reorientation back to its outward facing position.
Lipids
After digestion, the fats in our meal have been broken down into fatty acids, monoglycerides, and glycerol. They are absorbed primarily by simple diffusion of small particles across the brush border (the name for the microvilli-covered surface of the epithelial cells that line the small intestine) and by a small amount of active transport. The key here is the size of the fatty particles; they must be small in order to be absorbed. That’s where bile salts come in. The presence of a controlled flow of bile salts which break up the fats into tiny particles is essential for proper absorption of fats. If your gallbladder is not functioning properly or has been removed, you will have a problem absorbing fats. If you have a problem digesting fats for any reason, an option is to use ox bile tablets available at most health food stores. Supplemental digestive enzymes with lipase will also assist.
Another lipid of importance that is absorbed in the small intestine is cholesterol. As it turns out, cholesterol is readily absorbed in the small intestine. Specifically, a transport protein (NPC1L1) has been identified that transports cholesterol from the lumen (the interior space) of the small intestine into the enterocytes.
Note: unlike proteins and sugars, fats do not go directly into the bloodstream. They transport into the lacteals (tiny lymphatic ducts) found in the villi, and then travel through the lymphatic system and ultimately into the bloodstream. And in fact, fats do not enter the bloodstream in the form in which they were absorbed into the enterocyte. Once inside the enterocyte, fatty acids and monoglycerides are synthesized into triglycerides. These triglycerides are then packaged with cholesterol, lipoproteins, and other lipids into particles called chylomicrons. It is the chylomicrons that actually are transported into the lacteals and on into the bloodstream. Many doctors believe that a high triglyceride count in your bloodstream is actually more indicative of potential heart problems than a high cholesterol number.
Omentum
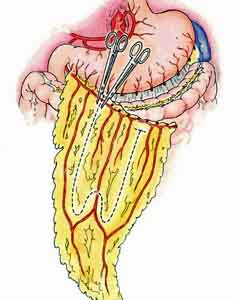 Okay, we need to revert to a little anatomy for a moment and talk about the omentum. It’s not really an organ, and it doesn’t really relate to digestion or absorption so it hasn’t made any sense to talk about it so far in our series on the intestinal tract. It does, however, relate to fat storage, and in that regard it makes sense to talk about it in terms of what happens to a large chunk of the fat we absorb.
Okay, we need to revert to a little anatomy for a moment and talk about the omentum. It’s not really an organ, and it doesn’t really relate to digestion or absorption so it hasn’t made any sense to talk about it so far in our series on the intestinal tract. It does, however, relate to fat storage, and in that regard it makes sense to talk about it in terms of what happens to a large chunk of the fat we absorb.
The omentum actually has two parts — the greater and the lesser. To keep things simple we’ll focus on the greater omentum, which hangs from the bottom of the stomach and extends down the abdominal cavity, then back up to the posterior abdominal wall after connecting with the transverse colon. The greater omentum is mostly made up of fat. It stores fat and provides a rich blood supply to the stomach. Specifically, it plays the following roles:
- It’s a fat depository, having varying amounts of adipose tissue. It’s one of your body’s primary storage sites for fat.
- Immune contribution, having milky spots of macrophage collections.
- Infection and wound isolation; It may also physically limit the spread of intraperitoneal infections. The greater omentum can often be found wrapped around areas of infection and trauma.
For the most part, these are “medical” considerations, but one aspect of the omentum will ring a bell for many readers. Sometimes when people lose a lot of weight, they wonder why their stomachs are still large and fatty. It’s often because of the fat stored in the omentum. The fat in the omentum is often the last fat to go when losing weight. If you want to lose the gut, you have to lose the fat from the omentum too.
Note: the lesser omentum is an attachment of the peritoneum that lies between the liver and the upper edge of the stomach. It carries the vessels that run to the stomach and liver.
Vitamins and Minerals
The thing to understand about vitamins and minerals is that for the most part, your body doesn’t like isolates, can’t absorb them, and considers them toxic if by chance they are absorbed. In general, your body prefers its vitamins and minerals bound to food — in their natural form, primarily bound to carbohydrates and some proteins. In fact, as might be guessed from all that we’ve learned about absorption in the small intestine, it’s actually the small lipids, sugars, and amino acids attached to the vitamins and minerals that the individual cells of your body recognize and absorb, not so much the vitamins and minerals themselves. Effectively, they just tag along for the ride into the cells. All that said, there are still important differences in how the different vitamins and minerals are absorbed.
Fat soluble vitamins
Assuming that your liver and gallbladder are working properly and that bile salts are breaking fats down into smaller, more absorbable particles, there is little problem absorbing the fat soluble vitamins — even when in an isolated form — such as d-alpha-tocopherol vitamin E. The bottom line is that the fat soluble vitamins (including vitamins A, beta- carotene, D, E and K) are diffused right along with their lipid carriers across the brush border of the cells found in the ileum. Likewise, they then travel with their associated fats on into the lymph system and then into the bloodstream.
The problem with using vitamin isolates when supplementing the fat soluble vitamins is not one of absorption or even one of toxicity (where the body thinks the isolate is a toxin). Rather, the problem is one of completeness. For example, consuming vitamin E as d-alpha-tocopherol leaves behind the seven other components of vitamin E (gamma, beta, and delta tocopherol — plus the four tocotrienols: alpha, beta, gamma, and delta). Likewise, supplementing with beta carotene or vitamin A leaves behind the several hundred other carotenoids that usually accompany them in nature — such as alpha carotene. Is that important?
Yes, very!
Studies have shown that alpha carotene is one of the most powerful carotenoids and has a strong inhibitory effect on the proliferation of various types of cancer cells such as those affecting the lungs, stomach, cervix, breast, bladder and mouth. It works by allowing normal cells to send growth-regulating signals to premalignant cells. Carrots, for that matter, contain approximately 400 different carotenoids in addition to beta carotene, and many of those carotenoids are far more powerful than beta carotene itself. If all you’re getting is beta carotene, you’re missing out. And if all you’re getting is synthetic beta carotene, you may actually be hurting yourself.
Water soluble vitamins
The water soluble vitamins such as vitamin C and most of the B vitamins are mainly absorbed in the jejunum. They are taken into receptor sites found on the villi by co-transport with sodium using the same transporter system used to carry monosaccharides into the bloodstream. These vitamins do present a problem when allowed to enter the bloodstream as isolates, no longer bound to their appropriate carbohydrates. First, by not being bound to the carbohydrates, it severely limits the amount of absorption that can take place (much of the supplement is wasted and passed on out through the rectum). Second, if absorbed in an isolated form, they are toxic to the body and are carried to the liver as “poisons.” The liver then neutralizes their toxicity through a process called conjugation that combines them with proteins. Although conjugation of water soluble vitamins stresses the liver (forcing it to do extra work), it does neutralize the toxic effect of the isolated water soluble vitamins and makes them usable by the cells of your body.
Minerals
 Minerals are absorbed in a small area at the top of the duodenum next to the pyloric valve where chyme passes out of the stomach. This is the primary absorption site for the bivalent minerals, including iron, calcium, magnesium, and zinc. The problem with minerals is that they are not easily absorbed in their raw isolated state (think oyster shells and iron filings) because of their electrical charge, which is opposite that of the intestinal wall. At first glance, this might seem like a good thing since opposite charges attract. Unfortunately, they attract to the extent that the minerals “stick” to the intestinal wall and do not get absorbed into the bloodstream. Eventually, the chyme moving through the intestinal tract pushes these “sticky” minerals down through the small intestine and on out through the rectum. Absorption of isolated minerals is about 3-5%. In a non isolated state, when bound to food, the charge is hidden, and absorption will be some ten times greater.
Minerals are absorbed in a small area at the top of the duodenum next to the pyloric valve where chyme passes out of the stomach. This is the primary absorption site for the bivalent minerals, including iron, calcium, magnesium, and zinc. The problem with minerals is that they are not easily absorbed in their raw isolated state (think oyster shells and iron filings) because of their electrical charge, which is opposite that of the intestinal wall. At first glance, this might seem like a good thing since opposite charges attract. Unfortunately, they attract to the extent that the minerals “stick” to the intestinal wall and do not get absorbed into the bloodstream. Eventually, the chyme moving through the intestinal tract pushes these “sticky” minerals down through the small intestine and on out through the rectum. Absorption of isolated minerals is about 3-5%. In a non isolated state, when bound to food, the charge is hidden, and absorption will be some ten times greater.
Manufacturers selling vitamin isolates, use a compromise. They chelate their minerals by wrapping amino acids around them. The amino acids “cover” the electrical charge and allow the minerals to be absorbed in the duodenum. Unfortunately, although the charge is obscured, isolates are not user friendly when it actually comes to utilization by the individual cells. In this case, absorption and utilization by individual cells are not the same thing and the rate of cell utilization is significantly less with chelated minerals. Food bound minerals, on the other hand, are easily absorbed through the small intestine AND they are readily utilized by every cell in the body.
An exception to this rule is what some marketers call “ionic minerals.” This is just a fancy way of saying that the mineral particles in the supplement (usually in a liquid form) are so small that the electric charge they generate is not strong enough to prevent its absorption. The bottom line is that good ionic mineral supplements (or their equivalent) are readily absorbed.
One other factor to consider is that the bivalent minerals are competitively absorbed because the area of absorption in the duodenum is relatively small. This means that an excessively high intake of one bivalent mineral in particular may occupy the entire absorption area and make the absorption of other bivalent minerals difficult. It also means that you need to supplement your minerals in an evenly balanced form rather than mega dosing on one mineral. To look at it another way, taking regular high doses of iron will impede the absorption of calcium, magnesium, and zinc leading to a series of other nutrition problems.
Proteolytic Enzymes
Many so called experts say that you cannot absorb proteolytic enzymes. First, they claim that as proteins, they are broken down by stomach acid and pepsin in the stomach unless they are enterically coated. Then other experts say that even if they did survive, their molecules are too big to pass through the walls of the small intestine. Whenever, I hear these arguments, I’m always reminded of the apocryphal story of the engineer who proved that bumblebees can’t fly. Applying the principles of aerodynamics, he PROVED that based on their size, weight, the size of their wings, and the physiological limits of how fast they could flap them, that bumblebees could not fly. Of course, how valid is a proof when the evidence before your eyes demonstrates it’s nonsense?
The absorption of proteolytic enzymes is a lot like the story of bumblebees. In the end, it doesn’t matter how many ways you try and prove that they can’t be absorbed; in the end, you can both measure them in the bloodstream and, more importantly, quantify the results of their presence in your own body.
In any case, let’s first deal with the digestive juice issue first. There are two rebuttals:
- Not all enzymes are destroyed by stomach acid and pepsin. Many are merely inactivated until they reach a friendlier pH environment such as found in the small intestine. Want an example of an enzyme that can survive stomach acid and digestive juices — in fact it thrives in a high acid environment? How about pepsin itself! Pepsin is an enzyme. Not only is it not destroyed by stomach acid; it’s actually activated by it. So much for the statement that all enzymes are destroyed in the stomach. (Really! Who are these people?)
- And even if all proteolytic enzymes were destroyed by digestive juices, instructions for using most such formulas tell you to take them between meals — when no digestive juices are present. Thus the issue is moot and the need for enteric coating moot…at least in a well designed formula used properly.
When I designed my own proteolytic formula, pHi-Zymes, I specifically selected enzymes that survive the stomach environment. It’s actually not that hard to do. The key is to use non-animal derived enzymes. Oral supplementation with non-animal derived enzymes, such as microbial enzymes — those manufactured by a fermentation process of Aspergillus, for example, possess unusually high stability and activity throughout a wide range of pH conditions (from a pH of 2-10). This enables them to be more consistently active and functional for a longer distance as they are transported through the digestive tract. Bottom line: they are not destroyed by stomach acid or pepsin.
Now let’s address the issue of absorption. The standard medical assumption is that no dietary protein is absorbed in an undigested form — pretty much without exception. Rather, since their molecules are too large, dietary proteins first must be digested into amino acids or di- and tripeptides before they can be absorbed. At first blush, that seems to exclude undigested enzymes (which are indeed proteins) from absorption. The clinker, though, is that enzymes, although they are proteins, are not dietary proteins. They are very different in function and structure; they are biochemical catalysts. In fact, enzyme molecules are much smaller than dietary proteins. In fact, they are smaller than DNA molecules. They are indeed small enough to be absorbed. The bottom line is that supplemental proteolytic enzymes can cross the intestinal wall.
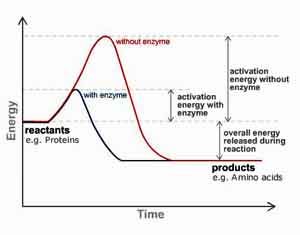
How exactly then are they transported across the mucosal membrane of the small intestine? The definitive answer appears to be unknown at this time. Nevertheless, studies indicate that proteolytic enzymes are able to increase the permeability of the mucosal epithelium and, hence, facilitate their own absorption by a mechanism of self-enhanced paracellular diffusion (i.e., across the tight junctions between the epithelial cells).
At this point, it’s probably worth abandoning our attempt to argue against the critics and return to the bumblebee analogy and examine what’s before our eyes. The bottom line is that if we can demonstrate that proteolytic enzymes consumed orally can later be found in the bloodstream, then we know they are absorbed no matter how many experts tell us they can’t get there — even if we don’t know exactly how they got there. And in fact, there are a plethora of studies that prove they reach the bloodstream.
- There are at least 17 studies that prove that nattokinase enters the bloodstream.
- Seaprose-S has at least six studies proving its efficacy on individuals with bronchial and sinus mucous as well as inflammatory issues.
- As for bromelain, there have been a number of studies over the years that substantiate its efficacy in the treatment of inflammatory disorders of the musculoskeletal system.
When summarizing the argument pro and con on the absorption of non-enterically coated proteolytic enzymes in the intestinal tract, I’m reminded of the movie Chicago. The husband of Kitty (Lucy Liu) says to his wife when caught in bed with two women, “Are you going to believe what you see or what I say?” In the end, it doesn’t matter what some experts say, proteolytic enzyme supplements can be seen in the bloodstream…and their benefits can be seen by anyone who uses them.
Fatigue after eating
And now let me touch on one final topic before concluding this newsletter on the absorption of nutrients in the small intestine: fatigue after eating. This appears to be one of those oxymorons that people have a hard time understanding. How can eating sometimes exhaust us?
We know that we can drink Gatorade or have a Snickers bar for quick energy in the middle of the day. But why is it that when we eat a larger, healthy, full spectrum meal (proteins, carbohydrates, and fats) that we actually feel enervated and sleepy for some time after eating, before the energy kicks in. And the answer is actually quite simple.
Digesting and absorbing food is energy intensive and exhausts the body. It takes energy for the body to produce stomach acid and pepsin. It takes energy for the body to produce the pancreatic enzymes that assist in digestion in the small intestine. And as we’ve seen in this newsletter, it takes energy to actually absorb proteins and carbohydrates across the enterocytes, into the villi, and on into the bloodstream. All in all, the body expends a great amount of energy getting nutrients into your bloodstream — enough energy so that you feel exhausted after eating a large meal. And the larger the meal, the more exhausted you feel. It is not until the digested/absorbed nutrients actually make their way through the bloodstream and on into every single cell in your body that you get your energy back. In the end, you gain more energy than you expended, and it is that energy that is used to power your body. But it can take several hours after eating to go from a negative expenditure of energy to a positive intake of energy and balance the scales out.
As a side note, taking supplemental digestive enzymes with your meals significantly decreases the fatigue factor experienced after eating large meals since they relieve your body of so much of its digestive work.

Conclusion
Okay, that concludes our exploration of the small intestine, both digestion and absorption. In our next newsletter we will pick up with the ileocecal valve, the gateway between the small and large intestines. From there we will explore how chyme (actually called fecal matter at this point) moves on through the large intestine and on out through the rectum. We will also explore all of the problems that can occur, including colorectal cancer and some of the options you have in dealing with them — both medical and alternative.










This didn't give me what I
This didn’t give me what I wanted,but it is a good place to find information.
Hello How much sodium (mgs)
Hello How much sodium (mgs) can one person digest within 24 hours that will not cause an imbalance and compromise ones nurtriton/health in both the small and large intestine? Regards, David
Sorry, there’s no simple
Sorry, there’s no simple answer to that. A healthy person, with plenty of potassium in the diet can easily handle upwards of 4,000 mg a day. For a person, with high blood pressure, compromised arterial function, and bad diet, even the low end recommendation of 1500 mg is too much. Also, the type of salt used matters.
I would like to ask you about
I would like to ask you about physiology of the small intestine in people which undergo total gastrectomy. Is any enzyme replacement useful for them?
Thank you
Answer:
Yes, digestive enzymes should be beneficial even after a gastrectomy. The more digestive enzymes present in your food when it reaches the intestinal tract, the less your body will have to produce to digest your food. This will take stress off your body and improve digestion – and nutrient uptake. That said, be sure and check with your doctor before incorporating enzymes as part of your daily routine.
I found your article when
I found your article when looking to review small intestine function/absorption since it’s been a while since physiology. Our doctor hypothesized that part of my wife’s immune response >> inflammation >> fibromyalgia could be in reaction to large proteins getting into the blood stream – “leaky gut” ; specifically casein, gluten, and soy protein. His analogy was that they aren’t digested properly and “punch holes” in the intestine wall…but since I knew most of what you describe here I wanted to see if there is a mechanism for this/does leaky gut make sense.
So, the point being, I was hoping to read a little more about paracellular diffusion and the possibility of large molecules/proteins making their way through somehow. Do you know anything about that, or those proteins in particular doing so? Would they somehow affect the tight junctions and enlarge the openings or….???
Thank you for the well-laid out info
Anything is possible, but we
Anything is possible, but we’ve never seen anything to validate the idea that proteins “punch holes” in the intestinal wall. Leaky gut, in fact, is usually associated with Candida overgrowth when the Candida cells transform into their fungal form and begin to grow long branches known as hyphae. It is these branches that then invade the cells in your intestinal lining and permeate the mucous membrane that prevents substances from leaking out.
That said, any allergen such as the proteins you mentioned is capable of eliciting an immune response and concomitant inflammation in cells that line the intestinal tract. If that inflammation gets severe enough, it can easily compromise the integrity of the intestinal wall.
What happened to excess amino
What happened to excess amino acids absorbed in small intestine?
Thank you so much! basically
Thank you so much! basically u save my life
i had a transvaginal done at
i had a transvaginal done at a teaching clinic ,and it was done wrong from the beginning,i had no idea of the problem, until 8 years later when i had an xray done of my torso,,,my small intestines were pushed to the sides both right and left ,i could not figure out why i was having so many digestion problems when i saw my xrays i asked the new doctor what is that,and he just looked at me and shook his head, can this be undone? can my small intestines be put back into place or not?